
Artículo:
Los dinosaurios terópodos son extremadamente raros en el Jurásico inferior y la mayoría de los informes son solamente de restos fragmentarios. Esta rareza se traduce en una diferencia considerable en nuestro conocimiento de estos animales en un momento determinado, todo parece indicar que los terópodos se diversificaron con rapidez. Los primeros terópodos reportados en Europa fueron en Escocia, Inglaterra, Francia y Bélgica y corresponden al Hettangiense, pero todos estos hallazgos son de material fragmentario.
The Oldest Jurassic Dinosaur: A Basal Neotheropod
from the Hettangian of Great Britain
- David M. Martill,
- Steven U. Vidovic,
- Cindy Howells,
- John R. Nudds
Los dinosaurios terópodos son extremadamente raros en el Jurásico inferior y la mayoría de los informes son solamente de restos fragmentarios. Esta rareza se traduce en una diferencia considerable en nuestro conocimiento de estos animales en un momento determinado, todo parece indicar que los terópodos se diversificaron con rapidez. Los primeros terópodos reportados en Europa fueron en Escocia, Inglaterra, Francia y Bélgica y corresponden al Hettangiense, pero todos estos hallazgos son de material fragmentario.

Imagen 1 Esquema del mapa que muestra Lavernock Point en el lado de Gales del estuario Severn.
Materiales, métodos y análisis utilizados
Los bloques más grandes se secaron durante dos semanas bajo
periódico húmedo para evitar la contracción rápida y agrietamiento de un fino
barniz de lutita en la superficie de piedra caliza. Los diversos laboratorios limpiaron
y prepararon mecánicamente para exponer los huesos usando un polvo abrasivo. Pequeñas fracturas de huesos
fueron reparados usando pegamento. Losas con los huesos representados como
moldes externos fueron fundidas en un compuesto de silicio, replicado en fibra
de vidrio y resina, y pintados para restaurar huesos que se sabe que han estado
presentes y se presumen perdidos en el mar. Existe la posibilidad de que parte
de la muestra permanece en el acantilado, pero la repetición de un examen
cuidadoso de la pared del acantilado no consiguió hallar nuevos huesos.
Para establecer la posición filogenética de la muestra se analizó
en un análisis cladístico compuesta de 366 caracteres y 46 taxones, de los
cuales 18 se encuentran fuera de Dinosauria.
Gracias a él análisis fue posible atomizar 25 de los caracteres
compuestos en la lista de caracteres de Ezcurra y Brusatte lo que resulta en
más de 28 caracteres. Un carácter compuesto adicional se redujo de tres estados
de caracteres a dos. Para demostrar el efecto de atomización caracteres
compuestos se llevó a cabo un experimento utilizando procedimientos jacknife (navaja
de bolsillo). El análisis de la navaja de bolsillo se realizó de forma manual
utilizando Microsoft Excel para generar caracteres aleatorios de omisión en
cada repetición del procedimiento.
El índice de retención de conjunto (PRI) que es
independiente del número de carácter no cambió entre el análisis original y el nuevo análisis, debido a que la codificación era exactamente la
misma. Del mismo modo, una gráfica de la diferencia entre longitud vs. el
número de caracteres eliminados demostró la misma velocidad de decaimiento de
longitud para ambas matrices. Sin embargo, la, nueva matriz más grande fue de
sólo 15 pasos, a pesar de otros 25 caracteres adicionales. Además, la
diferencia en la longitud relativa tomada de las líneas de regresión de la
trama jacknife fue de aproximadamente 60
pasos menos para la matriz más grande. Por lo tanto, la jacknife demuestra los
beneficios de la atomización de caracteres compuestos.
El análisis final se realizó en TNT mediante el uso de
"nueva tecnología". Se utilizaron pesos predeterminados para ajustar
los pesos de los caracteres durante el análisis, con el fin de combatir los
errores de codificación o conflictos de carácter que no se resolvió mediante la
atomización de caracteres compuestos. El MPT resultante se almacena en la
memoria RAM del ordenador se analizó adicionalmente con ATB intercambiando
algoritmo de búsqueda en "tradicional" de TNT para resolver los MPT
máximos posible.
¿Cómo datamos a este nuevo espécimen?
A pesar de que no se encuentra in situ, el nuevo dinosaurio está
contenido en uno de los dos horizontes de piedra caliza dentro de la formación
de Blue Lias inferior, y específicamente dentro de la parte baja del acantilado.
Por desgracia, este horizonte es una parte de la secuencia la cual no se ha
datado cronológicamente, son sitios por debajo de la primera aparición de la
amonita Psiloceras planorbis,
históricamente utilizado por estratígrafos británicos para definir la base del
Jurásico, y por encima de la última aparición de un marcador de conodontos del
Triásico, Chirodella verecunda Swift,
1995.
En las secciones europeas y norteamericanas en todo el
Triásico / Jurásico la primera aparición de la ammonites apsiloceratacean está marcada por la aparición de Psiloceruserugatum, seguido de P.imitansand P.antecedans, todos los
cuales se utilizan para definir subzonas debajo de la primera aparición de P.planorbis. En Austria P.spelaetirolicum, P.cf.pacificum y P.exgr.
P. tilmanni aparecieron en estratos por debajo de P.planorbis mientras que en Bélgica y noreste de Rusia, Primapsiloceras primulum se encontraron
en estratos por debajo de P. planorbis. Estas
subzonas del Jurásico temprano no han sido detectadas en la secuencia de
estratos. Sin embargo, se encontraron ammonites precediendo Psiloceras planorbis en la parte basal
de la formación Blue Lias a sólo 22 km
al sur de Lavernock Point .Estos
hallazgos incluyen ejemplos de P.
cf.erugatumand Neophyllites sp., lo que demuestra que los ammonites del Jurásico llegaban a las Islas Británicas.
La pregunta sigue siendo, si el nuevo dinosaurio atribuirse
al Triásico tardío, o al Jurásico temprano, el punto de vista geoquímico puede
ayudar a resolver la edad estratigráfica precisa del estrato en el que se
encontró el dinosaurio. Varios estudios isotópicos han situado el hallazgo
cerca del límite Triásico-Jurásico.

Imagen 2 Registro estratigráfico de Lavernock Point.
Un estudio realizado en Lavernock Point, encontró una
anomalía negativa δ13C org en la parte superior de Langport del grupo Triásico,
mientras que un análisis exhaustivo e integrado de una sección en el lado sur
del estuario de Severn en St Audries 'Bay, también detectó una anomalía
negativa de isótopos δ13C org que comienza justo por encima de las últimas
apariciones de conodontos, pero antes de la primera aparición de la amonita.
Esta excursión negativa también se ha detectado en otro lado, y en una sección
en el estado de Nueva York, EE.UU., Ward et al. Este análisis sirvió para
situar la base del Jurásico.
¿Triásico tardío o Jurásico temprano?
Situamos la base del Jurásico en los Alpes del Norte
austriacos. En esta localidad se define la base del Jurásico en la primera
aparición de los ammonites Psiloceras
spelae tirolicum. Estas especies de ammonites afloran aproximadamente 5 metros por encima de
la localización isotópica propuestas como la base del Jurásico.
Teniendo en cuenta que el nuevo dinosaurio se produce en una
posición estratigráfica por encima de la anomalía isotópica δ13 Corg; que se
produce por encima de un límite litológico claro que representa una facie
distinta que cambia de poca profundidad a la profundización de las condiciones
marinas; que se asocia con una fauna Jurásico, y analiza que el polen también
pertenece al Jurásico, el nuevo dinosaurio debe situarse a principios del
Hettangiense. Por lo que lo más probable es que pertenece a la subzona P.erugatum
del P.tilmanni , datada en unos 201,3
± 0.2million años.
Una vez resuelto el problema de su datación nos encontramos
con un nuevo problema, su situación filogenética.
Problema filogenético:

Imagen 3 Análisis cladístico de Dracoraptor hanigani.
Dracoraptor hanigani
posee 18 apomorfías, algunos de los cuales convergen con los artrópodos más
derivados, también conserva las condiciones plesiomórficas, tales como tres
dientes en el premaxilar.
Dracoraptor hanigani es
claramente un dinosaurio saurischian a causa de su construcción pélvica,
mientras que la posesión de dientes de sierra entre otros caracteres demuestra
sus semejanzas a los terópodos. El análisis cladístico sitúa a Dracoraptor dentro de Neotheropoda,
pero es basal en el clado. Dracoraptor se puede distinguir de Neotheropoda del Triásico tardío y Jurásico Temprano en una variedad de criterios,
no necesariamente apomórficos. Sin embargo, Dracoraptor
posee una combinación de caracteres basales que hacen que sea difícil colocarlo
filogenéticamente. La fosa antorbital superficial del maxilar y la presencia de
una muesca de obturador en el isquion indica afinidades Neotheropoda pero
muchos de las otros sinapomorfias Neotheropoda no pueden ser identificados en Dracoraptor.
Dracoraptor
conserva muchas características basales dentro de los artrópodos. Es probable
que las operaciones de preparación de la muestra permitan la identificación de
caracteres adicionales que pueden resolver mejor sus afinidades filogenéticas.
La presencia de Dracoraptor en estratos marinos puede ser de
poca importancia en relación con la ecología del animal, pero lo hemos
restaurado como un animal que habitaba en la línea de costa.
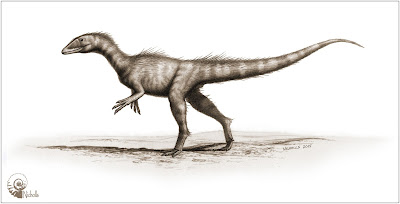
Imagen 4 Dracoraptor hanigani restaurado como una mezcla entre predador y carroñero.
Referencias
1.Benton MJ, Martill DM, Taylor MA. The first Lower Jurassic
dinosaur from Scotland: limb bone of a ceratosaur theropod from Skye. Scottish
Journal of Geology, 1995; 31:177–82. doi: 10.1144/sjg31020177
2.Owen R. II. Scelidosaurus harrisonii Owen of the Lower Lias. A monograph of
the fossil Reptilia of the Lias formations. Palaeontographical Society
Monographs, London. 1863; 14:1–26.
3.Woodward AS. Note on a megalosaurian tibia from the Lower Lias
of Wilmcote, Warwickshire. Annals and Magazine of Natural History, Series
8, 1908; 1:257–65. doi: 10.1080/00222930808692397
4.Andrews CW. On some remains of a theropodous dinosaur from the
Lower Lias of Barrow-on-Soar. Annals and Magazine of Natural History, Series
9 1921; 8:570–76.
doi: 10.1080/00222932108632620
5.Cuny G, Galton PM. Revision of the Airel theropod dinosaur from
the Triassic-Jurassic boundary (Normandy, France). Neues Jahrbuch für Geologie
und Paläontologie, Abhandlungen. 1993; 187:261–88.
6.Delsate D, Ezcurra MD. The first Early Jurassic (late
Hettangian) theropod dinosaur remains from the Grand Duchy of Luxembourg.
Geologica Belgica, 2014; 17:175–81.
7.Allain R, Tykoski R, Aquesbi N, Jalil N-E, Monbaron M, Russell
D, et al. An abelisauroid (Dinosauria: Theropoda) from the Early Jurassic of
the High Atlas Mountains, Morocco, and the radiation of ceratosaurs. Journal of
Vertebrate Paleontology, 2007; 27:610–24. doi:
10.1671/0272-4634(2007)27[610:aadtft]2.0.co;2
8.Hammer WR, Hickerson WJ. A crested theropod dinosaur from
Antarctica. Science. 1994; 264:828–30. pmid:17794724 doi:
10.1126/science.264.5160.828
9.Raath M.A. Morphological variation in small theropods and its meaning
in systematics: evidence from Syntarsus rhodesiensis;
pp. In: Carpenter K, Currie PJ. (eds.), Dinosaur Systematics: Approaches and
Perspectives. New York. Cambridge University Press, 1990; 91–105.
10.Rowe T. A new species of the theropod dinosaur Syntarsus from the Early Jurassic Kayenta
Formation of Arizona. Journal of Vertebrate Paleontology. 1989; 9:125–36. doi:
10.1080/02724634.1989.10011748
11.Talbot M. Podokesaurus holyokensis, a
new dinosaur of the Connecticut Valley: American Journal of
Science. 1911; 31:469–79. doi: 10.2475/ajs.s4-31.186.469
12.Welles SP. New Jurassic dinosaur from the Kayenta Formation of
Arizona. Bulletin of the Geological Society of America. 1954; 65:591–98. doi:
10.1130/0016-7606(1954)65[591:njdftk]2.0.co;2
13.Carrano MT, Hutchinson JH, Sampson SD. New information on Segisaurus
halli, a small theropod dinosaur from the Early Jurassic of
Arizona. Journal of Vertebrate Paleontology. 2005; 25:835–49. doi:
10.1671/0272-4634(2005)025[0835:niosha]2.0.co;2
14.Xu X, Zhao X, Clark JM. A new therizinosaur from the Lower
Jurassic Lower Lufeng Formation of Yunnan, China. Journal of Vertebrate
Paleontology. 2001; 21: 477–83. doi:
10.1671/0272-4634(2001)021[0477:antftl]2.0.co;2
15.Barrett P. The affinities of the enigmatic dinosaur Eshanosaurus
deguchiianus from the
Early Jurassic of Yunnan Province, People’s Republic of China. Palaeontology.
2009; 52:681–88. doi: 10.1111/j.1475-4983.2009.00887.x
16.Huene F. Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung
und Geschichte, Monographien zur Geologie und Palaeontologie. 1932; 1:1–361.
17.Tucker ME, and Burchette TP. Triassic dinosaur footprints from
south Wales: Their context and preservation. Palaeogeography,
Palaeoclimatology, Palaeoecology. 1977; 22:195–208. doi:
10.1016/0031-0182(77)90028-1
18.Kermack D. New prosauropod material from South Wales. Zoological
Journal of the Linnean Society, 1984; 82:101–17. doi:
10.1111/j.1096-3642.1984.tb00538.x
19.Yates AM. A new species of the primitive dinosaur Thecodontosaurus (Saurischia: Sauropodomorpha) and its
implications for the systematics of early dinosaurs. Journal of Systematic
Palaeontology. 2003; 1:1–42. doi: 10.1017/s1477201903001007
20.Storrs G. Tetrapods. In: Swift A, Martill DM. (eds) Fossils of
the Rhaetian Penarth Group. Palaeontological Association London, Field Guides
to Fossils Series. 1999; 9: 223–38.
21.Galton PM. Bones of large dinosaurs (Prosauropoda and
Stegosauria) from the Rhaetic Bone Bed (Upper Triassic) of Aust Cliff,
southwest England. Revue de Paléobiologie, Genève. 2005; 24:51–74.
22.Newton ET. On a megalosaurid jaw from Rhaetic beds near Bridgend
(Glamorganshire). Quarterly Journal of the Geological Society of London. 1899;
55:89–96. doi: 10.1144/gsl.jgs.1899.055.01-04.08
23.Galton PM, Yates AM, Kermack D. Pantydraco n. gen. for Thecodontosaurus
caducusYates, 2003, a basal sauropodomorph dinosaur from the Upper
Triassic or Lower Jurassic of South Wales, UK. Neues Jahrbuch für Geologie und
Paläontologie—Abhandlungen. 2007; 243:119–25. doi:
10.1127/0077-7749/2007/0243-0119



