
En algunas excavaciones los restos fósiles pueden contener
marcas que indican que el animal del que proceden no murió por causas
naturales. En esta entrada descubriremos cómo a partir de esas marcar podemos identificar el o los depredadores de la víctima en cuestión.

Nos centraremos
en un caso bastante reciente en el que las víctimas son Sphaerechinus granularis, un erizo de mar bastante común en el Mediterráneo. A la hora de identificar a la especie culpable, se ha de seguir un orden.
 |
| Sphaerechinus granularis camuflándose. |
1. Recogida de las pruebas: se recogieron muestras de equinoideos en las costas norte y occidental de la isla de Giglio, que se sitúa cerca de la costa occidental de Italia al norte del mar Tirreno (ver mapa debajo). En esta isla se pueden encontrar diversos hábitats.

Después
se procedió al secado de las muestras y a su medición. También se tomaron fotos
y se anotaron características tafonómicas como la presencia (o ausencia) de
tejidos blandos, rotura de espinas u otro tipo de partes, abrasión, etc.
2. Clasificación de las pruebas: se
recogieron 24 muestras, de las que 3 no mostraban ningún daño, 11 presentaban
perforaciones características de gasterópodos cásidos y 10 tenían evidencias de depredación durófaga.
3. Análisis de las pruebas: las 10 que
presentaban signos de depredación tenían diversos tipos de daño: orificios en
el lado aboral (A),
reducidos a la mitad del tamaño original (B) y restos pertenecientes a la zona
alrededor de la membrana peristomial (C). Se deduce que el daño siempre se produce, por tanto, desde la parte
contraria a la boca del equinodermo.


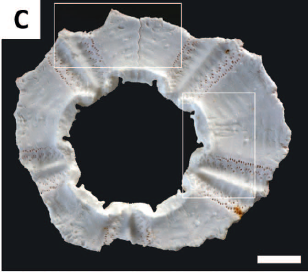
En el
interior a veces se podían encontrar arañazos y marcas producidas por dientes (D, E),
especialmente cerca de las heridas. Algunas muestras también presentaban
algunas espinas arrancadas (F).



Las muestras más recientes
estaban cubiertas de espinas y podían llegar a presentar restos de las partes
blandas (órganos). Cuanto más tiempo hubieran estado en el fondo marino, mayor
era el grado de incrustación de microorganismos (la muestra de la imagen B es más reciente que la de la imagen I).


4. Sospechosos: diversos organismos se
alimentan de Sphaerechinus granularis.
Los crustáceos usan sus pinzas para romper a su presa; los gasterópodos cásidos
usan ácido sulfúrico para penetrar al
interior; los asteroideos se comen a su presa y luego regurgitan los restos, o
digieren las partes blandas con estómagos extensibles; algunos peces usan
diversos métodos para romper a su presa; algunas especies de pájaros picotean dentro del
organismo o sueltan al equinoideo sobre una superficie dura para romperlos, y
las nutrias marinas usan piedras para romper al erizo de mar.
5. Identificación del culpable: el tipo de
rotura hace pensar que el culpable pertenece a los peces óseos. Estos peces
muerden primero las espinas y después ensanchan el agujero que se genera. Solo
unos pocos peces son depredadores de Sphaerechinus
granularis, ya sea porque ingieren individuos pequeños enteros o muerden y
rompen al erizo.
De
los organismos que se alimentan de este erizo de mar, solo los besugos alcanzan tamaños
que les permiten romper de esa forma a Sphaerechinus
granularis. Los más probables son Diplodus
sargus y Diplodus vulgaris, ambos
se pueden encontrar en las costas de Giglio. Las marcas de dientes encontradas en las
muestras se ajustaban a las dentaduras de ambas especies.
 |
| En la imagen de la izquierda se puede observar a Diplodus sargus en el centro y Diplodus vulgaris alrededor. |
Pero no siempre va a ser tan sencillo. Si vamos a realizar este mismo proceso pero con un fósil, nos encontraremos con algunos problemas extra. Para empezar, la preservación de marcas de depredación como las analizadas depende de varios factores, como la extensión y tipo de daño, el grado de incrustación de microorganismos (facilitan la conservación de las marcas) o la naturaleza del sustrato en el que se deposita el equinodermo. Hay un periodo crítico para la preservación de estas marcas: el proceso de incrustación y estabilización del resto se debe producir antes de que se degrade el tejido conectivo,si no se puede fragmentar y descomponer.
La potencial de preservación (es decir, la facilidad con la que se pueden conservar restos) depende de factores como la energía de las olas, la tasa de sedimentación y la ocurrencia de procesos atmosféricos puntuales (tormentas, por ejemplo). El caso en que se daría la mayor dificultad sería en aguas poco profundas que sufran mucha agitación y haya mucha abrasión.
En resumen, el estudio de la depredación nos puede permitir identificar organismos e incluso establecer relaciones entre ellos, lo cual es algo muy útil para establecer relaciones del pasado. Pero como hemos podido ver, es bastante complicado encontrar restos que tengan marcas, ya que no solo depende de que el resto las contenga sino también del ambiente donde se ha depositado el resto.

GLOSARIO:
.Cassidae: o cásidos, son moluscos gasterópodos con concha.
·Durofagia: comportamiento animal consistente en consumir organismos que tienen una concha dura o un exoesqueleto.
·Lado aboral: en asimetría radial, es el lado opuesto a la boca.
·Membrana peristomial: se encuentra en el lado oral y en su centro se encuentra la boca del equinidermo.
·Asteroideos: son una clase de organismos dentro de los equinodermos. Se conocen como estrellas de mar.
·Sparidae: familia de peces perciformes (con forma de perca), es decir, comprimidos lateralmente, aleta dorsal con espinas y aleta caudal escotada.
PARA SABER MÁS:
-https://es.wikipedia.org/wiki/Echinoidea
-https://www.ecured.cu/Asimetr%C3%ADa_radial
-https://practicasina2013.weebly.com/clase-echinoidea.html
REFERENCIAS:
-Diedrich Sievers & James H. Nebelsick. 2018. Fish predation on a Mediterranean echinoid: identification and preservation potential. Palaios, 33 (2): 47-54.
En resumen, el estudio de la depredación nos puede permitir identificar organismos e incluso establecer relaciones entre ellos, lo cual es algo muy útil para establecer relaciones del pasado. Pero como hemos podido ver, es bastante complicado encontrar restos que tengan marcas, ya que no solo depende de que el resto las contenga sino también del ambiente donde se ha depositado el resto.

GLOSARIO:
.Cassidae: o cásidos, son moluscos gasterópodos con concha.
·Durofagia: comportamiento animal consistente en consumir organismos que tienen una concha dura o un exoesqueleto.
·Lado aboral: en asimetría radial, es el lado opuesto a la boca.
·Membrana peristomial: se encuentra en el lado oral y en su centro se encuentra la boca del equinidermo.
·Asteroideos: son una clase de organismos dentro de los equinodermos. Se conocen como estrellas de mar.
·Sparidae: familia de peces perciformes (con forma de perca), es decir, comprimidos lateralmente, aleta dorsal con espinas y aleta caudal escotada.
PARA SABER MÁS:
-https://es.wikipedia.org/wiki/Echinoidea
-https://www.ecured.cu/Asimetr%C3%ADa_radial
-https://practicasina2013.weebly.com/clase-echinoidea.html
REFERENCIAS:
-Diedrich Sievers & James H. Nebelsick. 2018. Fish predation on a Mediterranean echinoid: identification and preservation potential. Palaios, 33 (2): 47-54.











{kind=link}