
Para continuar hablando de evolución por tierras lejanas, esta entrada la voy a
dedicar a la extinción del tigre de Tasmania y a la relación de la especie con
la evolución convergente. Esta vez me voy a basar en el estudio realizado por
Charles Y. Feigin, Axel H. Newton, Liliya Doronina, Jürgen Schmitz, Christy A.
Hipsley, Kieren J. Mitchell, Graham Gower, Bastien Llamas, Julien Soubrier,
Thomas N. Heider, Brandon R. Menzies, Alan Cooper, Rachel J. O’Neill y Andrew
J. Pask: "Genome of the Tasmanian tiger provides insights into the
evolution and demography of an extinct marsupial carnivor" (El genoma del
tigre de tasmania ofrece pistas sobre la evolución y demografía de un marsupial
carnívoro extinto).
El tigre de Tasmania (Thylacinus cynocephalus) fue la única especie dentro de la familia de Thylacinidae que sobrevivió hasta la era moderna. Era un marsupial carnívoro de tamaño grande que históricamente se distribuyó a lo largo de Australia antes de extinguirse en la zona hace alrededor de 3000 años. Esta extinción se dio por la manipulación ejercida por el ser humano en la especie.
Hará
unos 14000 años, la población de Tasmania se quedó aislada por un aumento en
los niveles del mar, hasta los comienzos del siglo XX. Los colonizadores europeos
vieron un gran efecto negativo de la especie sobre la ganadería ovina de
Tasmania, por lo que decidieron comenzar con la eliminación de estos animales.
El gobierno ofertó una paga de 1 dólar australiano por cada tilacino muerto. En
consecuencia, la especie decayó rápidamente en número, quedando oficialmente
extinta en 1982. El último tigre de Tasmania sabido murió en el Zoo de Hobart (Tasmania, Australia)
en 1936.

No obstante, lo que se estudia en el trabajo mencionado previamente, es la evolución convergente que esta especie ha tenido con los cánidos placentarios (o euterios). La evolución convergente es aquella en que dos especies con una filogenia completamente independiente (esto es, tienen distintos ancestros y, por tanto, distinta genética) terminan desarrollando estructuras físicas y de comportamiento similares de una forma 'curiosamente casual'.
El estudio
muestra cómo, sin tener un ancestro común, tanto los Tilacinos como los canidos
euterios han terminado evolucionando de forma muy similar. Esto viene explicado
en base a su similar alimentación carnívora en tiempos anteriores, que les ha
llevado a un igual desarrollo del cráneo y, por tanto, de muchas de sus formas
de actuar.
Este caso de evolución convergente es de los que más llaman la atención. La convergencia fenotípica es muy alta y, como el estudio ha demostrado, hay una parte genética que también converge, lo que se ha usado como explicación a la similitud de comportamientos.

Como dato, además, dado que la especie se extinguió, el foco de estudio se ha tornado hacia el llamado Demonio de Tasmania. Se ha investigado su posible relacion, en la evolución convergente, con otras especies. En el trabajo en que se basa esta entrada, el demonio de Tasmania es comparado, en concreto y en lo que a la evolución convergente respecta, con los euterios bóvidos; no obstante, la conclusión obtenida es de unos pocos genes convergentes (5 para ser exactos), y una clara no-convergencia evolutiva entre este demonio de Tasmania y los euterios bóvidos.
Os dejo una
foto del demonio de Tasmania, y algun enlace sobre ésta curiosa especie que se
ha convertido en el marsupial carnívoro más grande de Australia tras la
extinción del Tilacino.

Para terminar, dejo también el enlace a un blog que trata el tema (basándose también en una de las investigaciones que he utilizado como ayuda), y otros enlaces de interés:
https://kyletaitt.scienceblog.com/2013/02/07/the-curious-evolutionary-history-of-the-marsupial-wolf/
- Con respecto a la evolución convergente:
- Sobre el zoo de Hobart:
- Sobre los Euterios en general (enlace 1) y los bóvidos en particular (enlace 2):
- El Demonio de Tasmania (por qué ese nombre, historia e información básica):
REFERENCIAS
- Feigin, C. Y., Newton, A. H., Doronina, L., Schmitz, J., Hipsley, C. A., Mitchell, K. J., ... & Menzies, B. R. (2018). Genome of the Tasmanian tiger provides insights into the evolution and demography of an extinct marsupial carnivore. Nature ecology & evolution, 2(1), 182. ( https://www.nature.com/articles/s41559-017-0417-y )
- Paddle, R. (2002). The last Tasmanian tiger: the history and extinction of the thylacine. Cambridge University Press. ( https://books.google.es/books?id=wqIi6YTFPooC&printsec=frontcover&hl=es&source=gbs_ge_summary_r&cad=0#v=onepage&q&f=false )
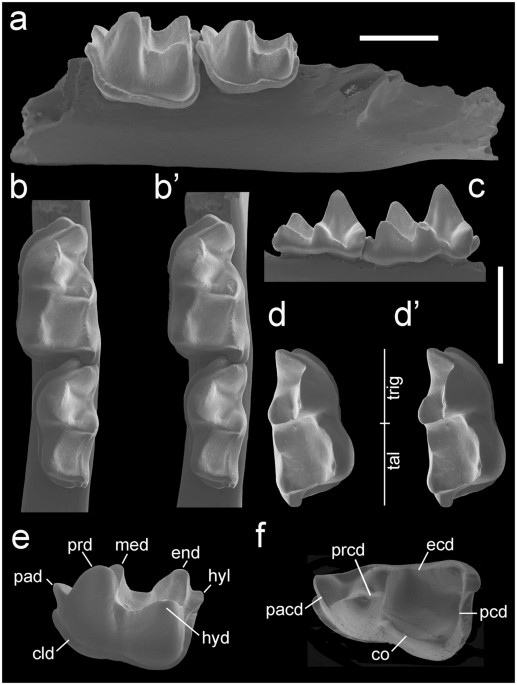
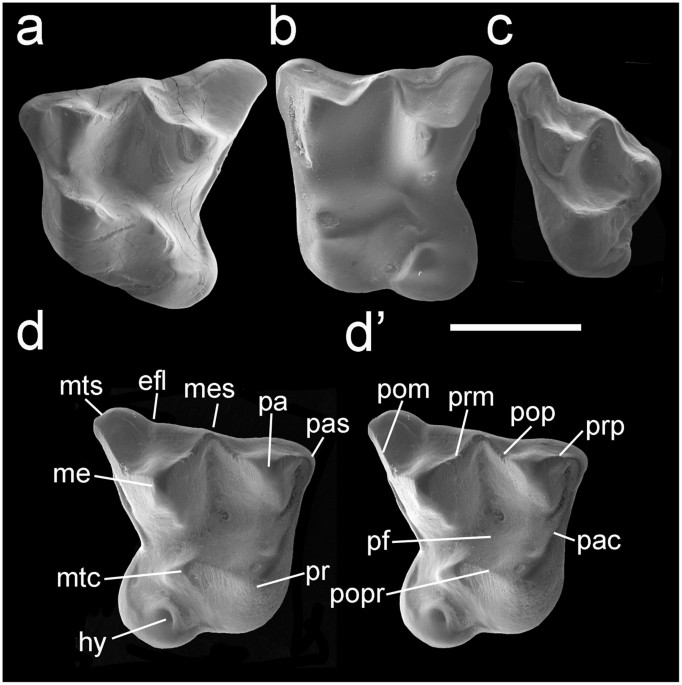
 † Indica taxón extincto; E, Early; L, Late; (#) , número de especies.
† Indica taxón extincto; E, Early; L, Late; (#) , número de especies. 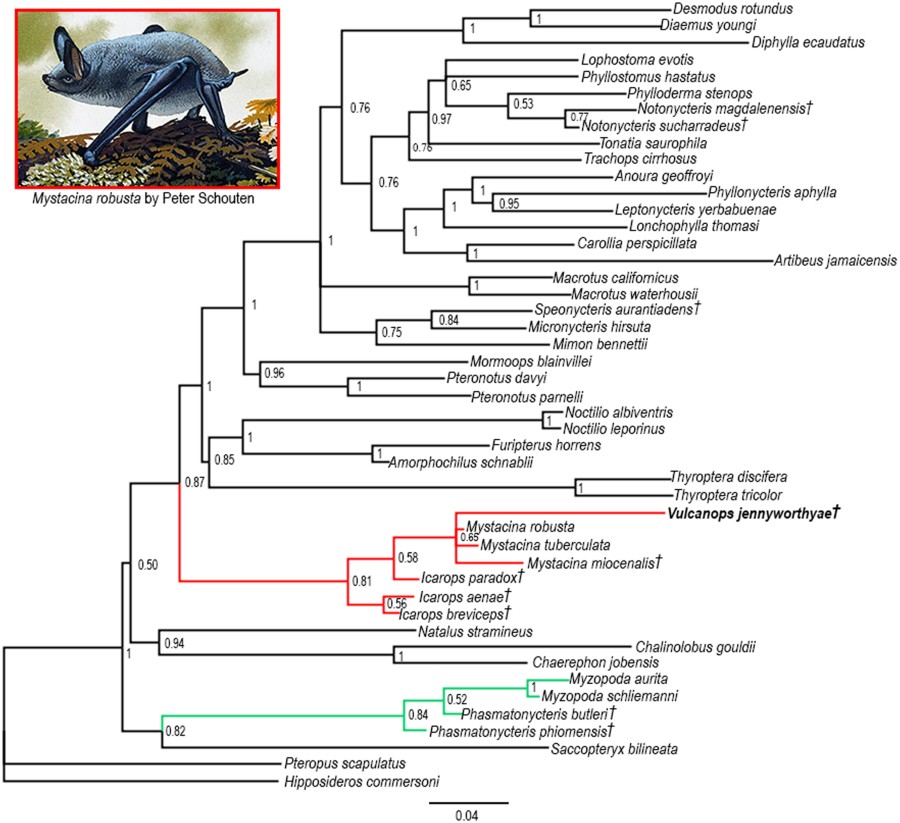

{kind=link}
{kind=link}
{kind=link}