

El otro día me topé con una noticia(os dejo el link de la noticia original por si os interesa) que decía que habían encontrado en el Cerro de los Batallones en Torrejón de Velasco (Madrid) restos de una nueva especie de mustélido. La verdad es que aquella palabra me sonaba y decidí buscar que era uno. Esta palabra hace referencia a animales mamíferos, carnívoros y fisípedos (pezuñas partidas). Se caracterizan por tener rostro y patas cortos, cuerpo alargado, garras muy afiladas, orejas pequeñas, pelaje bello y espeso y glándulas odoríferas. Algunos de estos son los hurones, las comadrejas o las martas. Lo cierto es que me dio curiosidad cuan parecidos podrían ser aquellos mustélidos a los que hay actualmente.
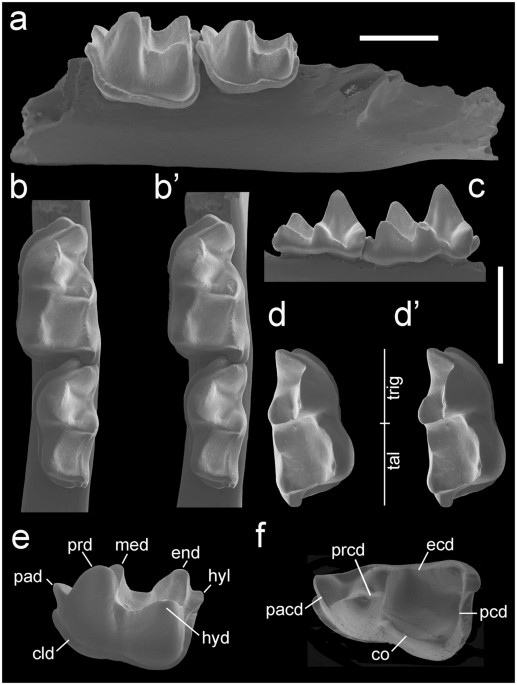
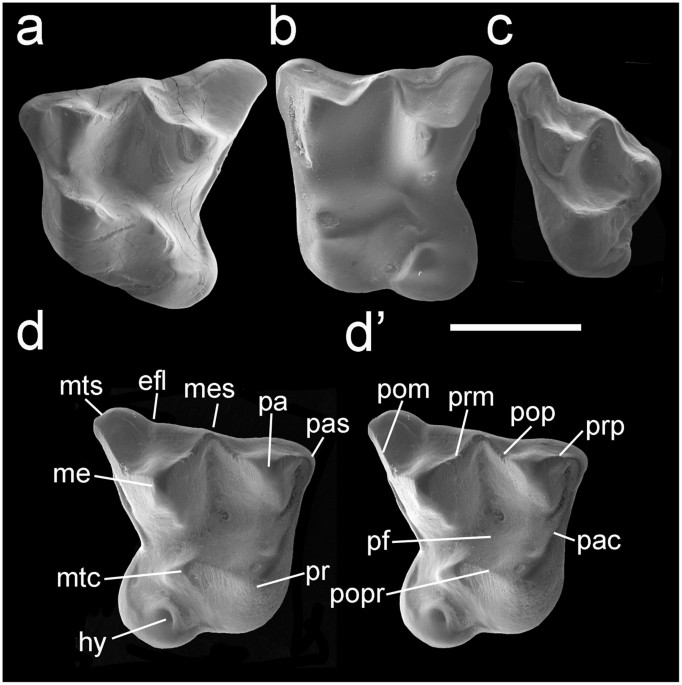
Al principio, tras el descubriendo de los restos fósiles en el yacimiento, pensaron que se trataba de una especie ya descrita, Circamustela dechaseauxi, pero tras la comparación del cráneo y restos dentales del ejemplar hallado, el de esta especie y otras especies de la misma familia pero pertenecientes al mismo u otro género se llegó a la conclusión de que se trataba de distintas especies y que los restos se trataban de una especie más primitiva. La nueva recibió el nombre de Circamustela peignei. La descripción de los fósiles encontrados se ha hecho a través del estudio directo (midiendo, observando y comprando las distintas piezas) y a través de técnicas no destructivas tomografía computerizada de rayos X (microCT-SCAN), con la que se pueden observar sitios que de forma convencional no se podría.

 mparación la Marta porque, irónicamente aunque se llame igual que yo, se parece mucho a esta especie y además es un animal que podemos encontrar en los bosques de España. La Marta vive en lo profundo de los bosques en pequeños espacios como huecos de árboles, piedras o, incluso, nidos realizados por otras especies. La Marta se puede alimentar de pájaros y sus huevos hasta el tamaño del urogallo, ardillas, lirones, ratas, ratones, reptiles, conejos y liebres, anfibios, aves de corral, huevos, frutos, miel, insectos y larvas, y hasta crías de corzo. Al tener una alimentación tan variada posee un total de 38 dientes, observándose en la mandíbula superior 6 incisivos, 2 colmillos, 8 premolares y 2 muelas, y en la inferior, 2 muelas más. En cambio, en en el caso de la Circamustela peignei, sus dientes tenían ciertas zonas más desarrolladas y mucho más punzantes y otras zonas, como la parte de los dientes que toca con la lengua, mucho más pequeña y menos desarrollada. Por lo que su mandibula sería mucho más primitiva, y como ya he mencionado antes, su alimentación sería más basada en carne de ahí que su dentición sea más afilada pues necesitaría desgarrar las piezas.
mparación la Marta porque, irónicamente aunque se llame igual que yo, se parece mucho a esta especie y además es un animal que podemos encontrar en los bosques de España. La Marta vive en lo profundo de los bosques en pequeños espacios como huecos de árboles, piedras o, incluso, nidos realizados por otras especies. La Marta se puede alimentar de pájaros y sus huevos hasta el tamaño del urogallo, ardillas, lirones, ratas, ratones, reptiles, conejos y liebres, anfibios, aves de corral, huevos, frutos, miel, insectos y larvas, y hasta crías de corzo. Al tener una alimentación tan variada posee un total de 38 dientes, observándose en la mandíbula superior 6 incisivos, 2 colmillos, 8 premolares y 2 muelas, y en la inferior, 2 muelas más. En cambio, en en el caso de la Circamustela peignei, sus dientes tenían ciertas zonas más desarrolladas y mucho más punzantes y otras zonas, como la parte de los dientes que toca con la lengua, mucho más pequeña y menos desarrollada. Por lo que su mandibula sería mucho más primitiva, y como ya he mencionado antes, su alimentación sería más basada en carne de ahí que su dentición sea más afilada pues necesitaría desgarrar las piezas.Estos nuevos hallazgos son de gran importancia ya que el Cerro de los Ballatones es uno de los yacimientos más importantes del mundo respecto a lo que se refiere a hallazgos de restos fósiles de mamíferos. De hecho se han encontrado restos de animales como cebras, tigres dientes de sable, rinocerontes..., en su mayoría carnívoros. Aunque estos descubrimientos demuestran que en el centro de la península también convivieron pequeños y medianos mamíferos y carnívoros que son también de gran interés para la comunidad científica.

Webgrafía
https://www.laaventuradelahistoria.es/cerro-de-los-batallones
https://www.clinicadentalsieiro.es/los-4-tipos-de-dientes-y-su-funcion/
https://www.hogarmania.com/mascotas/marta-mamifero-mustelido.html
https://www.faunaiberica.org/marta
https://www.clinicadentalsieiro.es/los-4-tipos-de-dientes-y-su-funcion/
https://www.hogarmania.com/mascotas/marta-mamifero-mustelido.html
https://www.faunaiberica.org/marta
Bibliografía
Valenciano.A., Pérez-Ramos.A., Abellla.J. & Morales.J. 2020. A new hypercarnivorous mustelid (Mammalia, Carnivora, Mustelidae) from Batallones, late Miocene (MN10), Torrejón de Velasco, Madrid, Spain. Geodiversitas 42(8): 103-121.





 † Indica taxón extincto; E, Early; L, Late; (#) , número de especies.
† Indica taxón extincto; E, Early; L, Late; (#) , número de especies. 

{kind=link}