
Foraminíferos desastres y oportunistas
en el Triásico temprano en el sur de China
HAIJUN SONG(1,2), JINNAN TONG(1),
PAUL B. WIGNALL(3), MAO LUO(4), LI TIAN(1) , HUYUE SONG(1), YUNFEI HUANG(5) & DAOLIANG CHU(1)
1-State Key Laboratory of Biogeology and
Environmental Geology, China University of Geosciences, Wuhan 430074, PR China
2-State Key Laboratory of Palaeobiology
and Stratigraphy, Nanjing Institute of Geology and Palaeontology,
Nanjing, 210008, PR China
3-School of Earth and Environment, University of Leeds,
Leeds LS2 9JT, UK [1]
4-School of Life and Environmental Sciences, Deakin
University, Melbourne Burwood Campus, Burwood, Victoria 3125, Australia
5-School of Geoscience, Yangtze University, Wuhan 430100, PR
China
Artículo: "Early Triassic disaster and opportunistic foraminifers in South China" y se ha publicado en la revista "Geological Magazine" / Volume 153 / Special Issue 02 / March 2016, pp 298-315
Artículo: "Early Triassic disaster and opportunistic foraminifers in South China" y se ha publicado en la revista "Geological Magazine" / Volume 153 / Special Issue 02 / March 2016, pp 298-315
Introducción
Para los sobrevivientes de las extinciones en masa, su
destino puede ser muy variable, pero también hasta cierto punto predecible. Los
grupos con altas tasas de extinción antes de la crisis a menudo irradian a
tasas elevadas, mientras que después rezagados evolutivos a menudo se recuperan
mucho más lentamente. Esto es ejemplificado por el proceso evolutivo de los ammonoideos,
un grupo caracterizado por excepcionalmente altas tasas de evolución a lo largo
de su historia.
Una de las facetas más interesantes del intervalo
post-extinción inmediata es la aparición de organismos oportunistas,
normalmente llamados taxones de desastres (Harries, Kauffman & Hansen,
1996; Kauffman & Harries, 1996).
Un total de nueve casos de especies oportunistas de foraminíferos
fueron identificadas de las 12 secciones del Triásico inferior en el sur de
China, Postcladella kalhori, Earlandia
sp., Globivalvulina lukachiensis, Hemigordiellina regularis, Hoyenella spp.,
Arenovidalina chialingchiangensis, Aulotortus? bakonyensis, Triadodiscus
eomesozoicus and Meandrospira pusilla.

Figura 1. La distribución estratigráfica de foraminíferos desastre yoportunista en las secciones inferiores del Triásico: Wufeng, Xiang Kou y Shangsi.
La distribución
temporal de los desastres y oportunistas
a) La distribución temporal de los desastres:
En este estudio, se encontró que los foraminíferos desastre
ocurrió en el período inmediatamente posterior a la extinción P-Tr (Fig. 2).
El grupo Postcladella Kalhori-Earlandia
sp., Por lo general dominado por abundantes Postcladellakalhori, Earlandiasp., Lukachiensis Globivalvulina y Nodosaria expolita, se produjo en los
estromatolitos en el Dajiang y secciones Cili que siguieron a la última
extinción masiva del Pérmico. Este grupo de foraminíferos también se han
encontrado en los estromatolitos basales del Triásico en la sección de Dongwan
del sur de China (Ezaki, Liu & Adachi, 2003) (Altineretal.1980;Altiner
& Zaninetti, 1981; Ünal et al. 2003; Groves, Altiner & Rettori, 2005).
Estos foraminíferos aparecieron con otros taxones desastre
como cianobacterias (Ezaki, Liu y Adachi, 2003; Wang et al., 2005), tubos de
gusanos poliquetos (Spirorbis) y
microgasterópodos (Yangetal.2011). Otro grupo de desastres dominado por Earlandia sp. ha sido identificado por
encima del horizonte de extinción del Triásico inferior. Varios picos en la
abundancia de Earlandia sp. se han
registrado coincidiendo con una extinción abrupta de foraminíferos durante la
crisis Triásico temprano.


Figura 3. Número de muestras en función del número de géneros en una sección delgada de 2,2 x 2,2 cm2 en el caso de desastres oportunista desde el Triásico Inferior y taxones normales desde el Pérmico superior y el Triásico Medio.
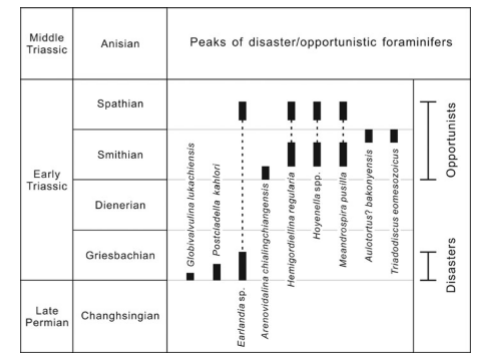
Figura 2. Rangos estratigráficos foraminíferos desastre y oportunista en el sur de China durante el final del Pérmico y el Triásico temprano.

b) La distribución temporal de los oportunistas:
En los estratos Dieneriense del sur de China, no encontramos
ningún foraminífero oportunista o de desastre. La fauna oportunista domina
dentro de la de Smith y Spathian y se caracteriza por la gran prosperidad Hemigordiellina y Hoyenella (Fig. 2). El número de especímenes de Hemigordiellina y Hoyenella spp. supera 200 en una lámina delgada 2,2 × 2,2 cm2 en
algunos niveles en la sección Shangsi (Fig. 3).
El límite inferior del intervalo de la fauna oportunista se
define por el horizonte, donde Arenovidalina
chialingchiangensis apareció por primera vez. El límite superior del
intervalo de la fauna oportunista se define por el horizonte, donde la
abundancia relativa del grupo oportunista disminuye a menos del 50%.
Los portunistas smithian se dividen en tres grupos en
función de las gamas estratigráficas. El primer grupo dominado por Arenovidalina
chialingchiangensis se produjo en la Formación Tongjiez inferior (Fig. 1). El
segundo grupo dominado por Hemigordiellina
regularis, Hoyenella spp. y Meandrospira pusilla se produjo en la
Formación Tongjiezi medio (Fig. 1). El tercer grupo dominado por Aulotortus? Bakonyensis y Triadodiscus eomesozoicus ocurrió en la
Formación Tongjiezi superior (Fig. 1). La fauna oportunistas en el Spathian
consisten en Earlandia sp., regularis
Hemigordiellina, Hoyenella spp. y
Meandrospira pusilla (Fig. 2).
Estrategia de
supervivencia como respuesta a los ambientes de estrés
Los taxones oportunistas por lo general se aprovechan de
situaciones de alto estrés, generando fuertes ambientes fluctuantes como
resultado de los cambios dramáticos en los ecosistemas oceánicos. Como tales,
son capaces proliferar y producir una expansión de la población C y una rápida
dispersión biogeográfica en ambientes estresados (Harries, Kauffman y Hansen,
1996; Kauffman y Harries, 1996).
En este estudio, se encontró que los foraminíferos desastres
adquieran poblaciones relativamente
grandes en el intervalo de supervivencia temprana. Son reemplazadas por
foraminíferos oportunista y otros sobrevivientes de los primeros en el
siguiente período de repoblación. Ambos tuvieron un lapso de tiempo muy corto,
y aparecieron en varias ocasiones en el Triásico inferior. Estas camas fósiles
por lo general tienen una baja diversidad pero una alta abundancia (Fig. 3).
Las especies 'normales' (incluyendo los foraminíferos
Pérmico y Triásico tardío que se han reportado en Song en 2007, 2009, 2011,
2015;. Song, Tong y Chen, 2009, 2011, y algunos no publicados para el Dongling,
Meshikou. , secciones Coli, y Bianyang) por lo general tienen una diversidad moderada
con una abundancia moderada (ver Fig. 3). Estos taxones oportunistas y
desastres son muy pequeñas en comparación con las formas pre-extinción (Payne
et al 2011;. Song, Tong y Chen, 2011; Regoet al 2.012.). Todos estos rasgos
caracterizan a una estrategia de selección de r, es decir, alta fecundidad, tamaño
corporal pequeño, corto tiempo de generación y dispersión y amplia
descendencia. Cuando las condiciones ambientales tienden a mejorar los K
estrategas empezaron a dominar el ecosistema bentónico.
Conclusión
Tres especies de foraminíferos desastre fueron identificados
en el período inmediatamente posterior a la extinción en masa P-Tr, i.e.Postcladellakalhori, Earlandia sp. y Globivalvulina lukachiensis. Entre
ellos, Postcladella Kalhori y Earlandia sp. También se ha
encontrado que las especies de desastre en muchas otras regiones del mundo.
Como tal, la floración (en lugar de la ocurrencia) de estas formas de desastre
podrían ser utilizados como evidencia de los estratos posterior extinción en el
caso de una falta de conodontos y amonoides.
La fauna desastre fue sustituidos por la fauna oportunista
en Smithian y Spathian .La fauna oportunista está compuesto por Earlandia sp., Hemigordiellina regularis, Hoyenella spp., Arenovidalina chialingchiangensis, Aulotortus? bakonyensis,
Triadodiscuseomesozoicus y Meandrospira
pusilla. Esta fauna oportunista son los principales componentes de la fauna
de recuperación (véase Song et al. 2011), y la abundancia relativa disminuye a
menos del 50% en el Spathian medio alto,
de acuerdo con la mejora de los ambientes marinos.
Foraminíferos oportunistas y desastres a menudo se han
encontrado en las secuelas de muchos eventos de extinción en el Fanerozoico.
Este fenómeno muestra r-selección, una estrategia usada comúnmente por
sobrevivientes para hacer frente a los eventos catastróficos. Después de la
crisis, muchos oportunistas viven en áreas limitadas, mientras que otros son
propensos a elegir una estrategia de selección de K y se convierten en los
grupos dominantes durante el intervalo de recuperación.
Referencias
ALTINER,D.&ZANINETTI,L.1981.Le Triasdans la régionde
Pinarbasi, Taurusoriental, Turquie: unités lithologiques, micropaleontologie,
milieux de dépôt. Rivista Italiana di Paleontologia 86, 705–60.
EZAKI, Y., LIU, J. & ADACHI, N. 2003. Earliest Triassic
microbialite micro- to megastructures in the Huaying area of Sichuan Province,
South China: implications for the nature of oceanic conditions after the
end-Permian extinction. Palaios 18, 388–402.
EZAKI,Y.,LIU,J.,NAGANO,T.&ADACHI,N.2008.Geobiological
aspects of the earliest Triassic microbialites along
thesouthernperipheryofthetropicalYangtzeplatform: initiation and cessation of a
microbial regime. Palaios 23, 356–69.
GROVES, J. R., ALTINER, D. & RETTORI, R. 2005.
Extinction, survival, and recovery of lagenide foraminifers in the
Permian–Triassic boundary interval, Central Taurides, Turkey. Journal of
Paleontology 79(suppl.), 1–38.
HARRIES, P. J., KAUFFMAN, E. G. & HANSEN, T. A. 1996.
Models for biotic survival following mass extinction. In Biotic Recovery from
Mass Extinction Events (ed. M. B. Hart). pp. 41–60. Geological Society of
London, Special Publication no. 102
KAUFFMAN, E. G. & HARRIES, P. J. 1996. The importance of
crisis progenitors in recovery from mass extinction. In Biotic Recovery from
Mass Extinction Events (ed. M. B. Hart), pp. 15–39. Geological Society of
London, Special Publication no. 102.
1 comentario:
Por sí a alguien le interesa, el artículo se titula "Early Triassic disaster and opportunistic foraminifers in South China" y se ha publicado en la revista "Geological Magazine" / Volume 153 / Special Issue 02 / March 2016, pp 298-315
Publicar un comentario