 Nieves López y Jaime Truyols en su obra Paleontología
(
Nieves López y Jaime Truyols en su obra Paleontología
(López Martínez & Truyols Santonja (1994)
Paleontología. Síntesis,
Madrid) sostienen que
“La
Paleontología no sólo se ocupa del estudio de los organismos del pasado en sí
mismos, sino también de los cambios que se han ido produciendo en el mundo
biológico de manera global a través de los tiempos”.
Tomando esta definición como excusa, y la temática de las
momias que dejamos en el ultimo post
como hilo conductor, me propongo a tratar en esta ocasión un tema que los más
ortodoxos considerarán, a priori , fuera
del campo de la Paleontología: las momias humanas, y lo que nos han enseñado;
aplicado obviamente al conocimiento de las formas de vida en el pasado, de la
paleobiogeografía, la arqueobotánica, la paleoclimática o paleoambiente, la paleoparasitología o la arqueomedicina, entre otros muchos campos de la Biología
Es cierto que el estudio de las momias humanas se lo tenemos
que dejar a los arqueólogos, pues de eso son expertos; pero el hecho de que los
restos humanos estén momificados (natural o artificialmente) hace necesario en
el estudio de cualquier hallazgo de este tipo la colaboración de médicos o químicos
entre otros muchos expertos: en ese momento entran en juego los Paleontólogos.
Primera parada: El sur del Tirol: Ötzi.
 |
| Gorro. Ursus arctos |
Y es que ¡de restos humanos
momificados se puede sacar una cantidad de información E. Egarter Vigl et al. 2001); y que en el año 1991 fue encontrado tal cual murió, en un
glaciar del sur del Tirol, en la frontera entre Austria e Italia. Todo tipo de
estudios se han llevado a cabo en Ötzi, por ejemplo, se sabe gracias a sus
ropas que en esa época los osos (Ursus
arctos arctos) eran abundantes en los Alpes, pues lleva prendas de abrigo
hechas de piel de oso, principalmente el gorro. También se ha podido comprobar
que sus congéneres tenían rebaños de cabras domesticadas, y que usaban ese
cuero para confeccionar el calzado, por ejemplo. Se ha analizado también su
contenido intestinal (Oeggl K et al., 2006.) para hallar que sus dos últimas comidas fueron ciervo (Cervus elaphus) y rebeco o chamois(en francés,
la subespecie de la zona) (Rupicapra r. rupicapra);
aunque lo más interesante de lo encontrado en su estomago, y lo que más pistas
ha dado acerca del cadáver ha sido el polen en su tracto digestivo y el resto
de su cuerpo. Según el trabajo del paleo-botánico Klaus Oeggl se ha podido,
a través de los pólenes hallados, establecer que la muerte se produjo en
primavera -entre los meses de abril o mayo-, o que su última comida fue en un
bosque de coníferas…¡ y hasta se especula que hacía viento durante esa comida!
 |
| Calzado de Ötzi, elaborado con fibras vegetales. |
increíble! Por ejemplo: el
caso del famoso Hombre de Ötzi; un adulto de la Edad de Cobre;( hace
aproximadamente 5.300-5.600 años) que tuvo la brillante idea de tomar un atajo
en su camino, de vuelta de comer… y murió a consecuencia de una supuesta
emboscada (
Estas y otras muchas sorpresas son las que nos ha regalado
el estudio de Ötzi, en concreto el aporte de expertos en todas la áreas de la
Biología. Los mayores hitos acerca de la investigación en la momia de Ötzi las
podéis encontrar en
esta página del Museo Arqueológico del sur del Tirol, (y
recomiendo aunque sea una visita rápida para ver la cantidad de información que
se le ha podido sonsacar, gracias a la tecnología actual, a este magnífico
descubrimiento.)
Del frío alpino a la brisa cálida del Nilo.
Nuestros
siguientes sujetos de estudio no necesitan introducción. El concepto de momia se asocia inconscientemente, quizá
por la fascinación casi infantil que en muchos de nosotros provocan, a las
formas de enterramiento típicas del antiguo Egipto, que son famosas por las
románticas figuras de los varios Howard Carters:
buscatesoros, arqueólogos, anticuarios, historiadores… egiptólogos en resumen;
un campo efervescente en los últimos tres siglos, pero dejado un poco de lado a
mediados del siglo pasado. Hasta que, como en casi todo campo del saber
empírico, la revolución técnico-científica que vivimos actualmente abrió la
puerta a respuestas que jamás los clásicos egiptólogos hubieran buscado.
El análisis genético y bioquímico de las momias y sus
ajuares funerarios llevó al campo de la egiptología a bioquímicos, botánicos,
entomólogos, e incluso a investigadores de la evolución y la filogenética. Y es
que las momias egipcias tienen la particularidad de tratarse de momias
artificiales: preparadas por el hombre, es decir, que contienen en ellas restos
de las preparaciones utilizadas en el proceso de momificación: una puerta de
acceso a la medicina y la farmacia de la época.
 |
| Erythroxylum coca, la planta de Coca. |
En este contexto, la botánica Dominique Görlitz, en el año
2016 se atrevió a publicar un trabajo que destruyó las bases del paradigma
histórico en el que siempre se han basado las historias paralelas del “Viejo” y
“Nuevo” Mundo, antes del 1492. En “The Occurrence of Cocaine in Egyptian
Mummies -New research provides strong evidence for a trans-Atlantic dispersal
by humans”, Dominique afirma haber encontrado trazas de Nicotina y de
Cocaína en un análisis bioquímico de varias momias encontradas en el Valle de
los Reyes, en Egipto. La
botánica señala que “one
could still try to explain the presence of tobacco in ancient Egypt with local,
previously unknown African species of tobacco, but with cocaine it is not
possible”. Y es que, aunque existe alguna especie del género Erythroxylum en África, sólo cuatro especies de este
género presentan los alcaloides que componen la cocaína (entre ellas Erythroxylum
coca la más extendida), y sólo se encuentran, y se cultivan, en las
primeras estibaciones de los Andes, donde todavía domina el paisaje de la selva
tropical.
Si lo que postula D.
Görlitz es cierto, implica que hubo encuentros culturales, y seguramente hasta relaciones
comerciales entre los pueblos originarios sudamericanos y la antigua
civilización egipcia: globalización en
el año 1000 a.C., aproximadamente.
Las primeras y las últimas momias: las más desconocidas.
Un
último ejemplo de implicación de paleontólogos en los estudios de momias nos
lleva a las momias Incas. Estas últimas son muy desconocidas, aunque derivan de
una larga tradición que tenían los pueblos precolombinos, habitantes de las
zonas más áridas de Sudamérica, de enterramientos en estilo momia. De hecho, las momias humanas encontradas más antiguas
del mundo; fruto de momificación artificial, son precisamente las halladas en
el cerro de Arica, en el extremo norte de Chile, pleno desierto de Atacama:
datan de hace 9000 años. (Guhl, F. et al., 2000)
Los incas, en su etapa de esplendor, dejaron, desperdigados
por los Apu de la Cordillera de los Andes, las montañas más importantes del Tawantinsuyu –del
imperio- , vestigios de la ceremonia del Capac Cocha, que consistía entre otras
cosas de dejar a uno o varios jóvenes entumecidos de frío en una posición
semienterrada en la cima de cualquiera de estas montañas como ofrenda a su dios
Viracocha. Estos sacrificios humanos, por efecto del clima de alta montaña, y
la sequedad del desierto, se momificaban naturalmente, por el mismo proceso que
el hombre de Ötzi: por liofilización. Estas momias han sido las madres de mucha
de la información que tenemos a día de hoy del estilo de vida del pueblo Inca,
ya que no tenían ningún tipo de lenguaje escrito en el que pudieran plasmar su
historia y vida.
 |
El Niño del Plomo, en exposición en el Museo
de Historia Natural de Santiago de Chile. |
A día de hoy, hay muchos estudios de Paleoparasitología, es decir,
paleontología de los patógenos, que se han interesado en el estudio de estas
momias, pues muchas de ellas son de la época de la conquista española del continente,
y las momias son capaces de mostrar la velocidad de propagación de las plagas o
enfermedades traídas por los conquistadores, entre otras muchas cosas.
Quiero destacar de este apartado dos tipos de sujetos investigación,
pues son muy abundantes las momias de animales, especialmente de Cuys o
Conejillos de indias (Cavia porcellus), codiciado alimento entre las comunidades andinas de
tiempos inmemoriales, y que se sigue consumiendo en la actualidad. En estos
restos animales se estudia la presencia de exo y endoparásitos, y su relación
que pueden tener con los españoles, para poder establecer patrones de
dispersión de los parásitos que puedan ser aplicables en la actualidad. (Dittmar, K., et al. 2003) (Ewing, H.E., 1924)
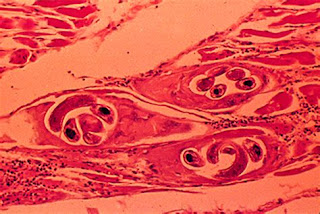 |
Quistes producidos por la Triquinosis en el músculo esquelético.
Tinción: Hematoxicilina-Eosina; así se diagnosticó inicialmente la
enfermedad al Nño del Plomo. |
En cuanto a enfermedades humanas, el ejemplo más interesante
o controversial (para seguir con la línea conspiratoria
del trabajo de Dominique Görlitz), es el de la Momia del Niño del Plomo, una
momia del Capac Cocha encontrada en el Cerro del Plomo ( 5.200-5.400 m.s.n.m.),
montaña visible desde la cuenca de Santiago, en la que se emplaza la capital de
Chile. En la momia de este niño, de unos nueve años de edad, científicos de la
Universidad de Chile diagnosticaron, en el año 2010, triquinosis (Rodríguez, Héctor, et al., 2011); una
enfermedad causada por la ingesta de carne
con larvas de nematodos del género Trichinella
sp.y que se sabe que era inexistente en América antes de la llegada de los
europeos. El dilema se encuentra en que la muerte del Niño del Plomo está
datada alrededor del año 1500, 37 años antes de que se cualquier europeo se
acercara siquiera al actual territorio chileno. Sin el estudio de más muestras
humanas y animales de esta época no se puede establecer si la enfermedad
también existía en América, o si, como los animales domésticos actúan como vector
de transmisión de esta enfermedad, no hizo falta que los conquistadores
llegaran a entrar en contacto con los Incas para que estos ya empezaran a
padecer la enfermedad: una dura premonición, (transmitida por nematodos) de lo que se les venía encima…
Concluyo así, que el estudio de las momias humanas por parte de expertos paleontólogos no puede traer más que sorprendentes descubrimientos, y se trata de una mirada profesional que no se le ha dado a la mayoría de las momias en tiempos anteriores.
Bibliografía.
- López Martínez & Truyols Santonja (1994) Paleontología. Síntesis, Madrid.
- Murphy, William A., et al. “The Iceman: Discovery and Imaging.” Radiology, vol. 226, no. 3, 2003, pp. 614–629.
- Oeggl, Klaus, et al. “The Reconstruction of the Last Itinerary of ‘Ötzi’, the Neolithic Iceman, by Pollen Analyses from Sequentially Sampled Gut Extracts.” Quaternary Science Reviews, vol. 26, no. 7-8, 2007, pp. 853–861.
- Görlitz, Dominique. The Occurrence of Cocaine in Egyptian Mummies New Research ... Technische Univerität Dresden, Institut Für Kartographie, Germany. 2016.
- Guhl, Felipe et al. Chagas disease and human migration. Mem. Inst. Oswaldo Cruz[online]. 2000, vol.95, n.4 [cited 2018-03-20], pp.553-555.
- Dittmar, K, et al. “Techniques of DNA-Studies on Prehispanic Ectoparasites (Pulex Sp., Pulicidae, Siphonaptera) from Animal Mummies of the Chiribaya Culture, Southern Peru.” Memórias Do Instituto Oswaldo Cruz, vol. 98, no. suppl 1, 2003, pp. 53–58.
- Ewing HE 1924. Lice from human mummies. Science 60: 389-390.
- Rodríguez, Héctor, et al. “Análisis Paleoparasitológico De La Musculatura Esquelética De La Momia Del Cerro El Plomo, Chile: Trichinella Sp.” Chungará (Arica), vol. 43, no. especial, 2011, pp. 581–588.
















{kind=link}
{kind=link}
{kind=link}