
assemblage from the Lower
Cretaceous Jiaguan Formation in the
Sichuan Basin, southwestern China:
A new ornithischian ichnotaxon,

Hola de nuevo amigos! Como os habéis podido dar cuenta, mis dos entradas anteriores se han centrado en la lomoción de los dinosaurios, y en esta nueva entrada también os voy a hablar de ello. Con cada entrada intento atraeros al apasionante mundo de la icnología, y transmitiros lo asombroso que es saber qué organismo originó la huella y cómo era su forma de andar. Intento no ser pesada pero no puedo evitar dejar de hablar de ellas debido a mi gran afición. Espero que os guste esta nueva entrada, y se os haga entretenida la lectura.
 |
| Figura 1. Cuenca de Sichuan. |
Nos adentramos en China. En concreto en la cuenca de Sichuan, conocida también con el nombre de cuenca Púrpura o Roja.
Es una de las cuencas más grandes de China, forma parte de la plataforma de
Yangtze, y contiene materiales datados
del Mesozoico. En su margen meridional, donde se encuentra ubicado Shimiaoguo, y se han encontrado con un área de huellas en varios bloques caídos, pertenecientes a la
formación Jiaguan (Xing et. al, 2015).
 |
| Figura 2. Mapa que representa la provincia de Sichuan, señalando con una estrella los lugares donde se han encontrado huellas de dinosaurios.
Cada camino de huellas lo numeraron con el prefijo SMG, y fueron realizadas por terópodos, designados con la mayúscula T; D para dromeosáuridos; S para saurópodos; O para ornitópodos; P para pterosaurios (Zhen et.al 1994), (Xing et.al 2013), (Xing et.al 2015). Lo primeron que hicieron fue realizar una tabla con varias columnas donde represetaron cada uno de los bloques, el organismo que las originó (th: terópodos no voladores; Dr: dromeosaurios; Sa: saurópodos; Or; ornitópodos; Pt: pterosaurios), y la relación entre el número de huellas (TS) y el rastro dejado (TWS).
RECOPILACIÓN DE DATOS. Tras una intensa y dura recogida de datos, se realizaron una serie de tablas:
EL PRIMER ESTUDIO: HUELLAS DE TERÓPODOS.
Comenzaron analizando las huellas de terópodos encontradas en los 8 bloques, excepto en el bloque 7 que no encontraron ninguna. Han podido identificar 31 pequeñas huellas tridáctilas cuya longitud del pes era de 7-18 cm, y las clasificaron en 7 caminos diferentes catalogadas como T1-T7, gracias a la ayuda de toda la información recopilada en la tabla 2. (Figura 3).
|
 |
| Figura 3. En el bloque 1 aparecen diferentes impresiones de huellas realizadas por tres grupos diferentes de organismos: terópodos, saurópodos y ornitópodos, siendo los predominantes los terópodos. |
Se pudo saber con exactitud que eran de terópodos gracias a la mejor huella conservada, que aparece en la figura 4. En esta huella se aprecia claramente como la longitud del dígito III es la más larga y los dígitos exteriores son casi iguales, el ángulo entre los dígitos II-IV es de 76º (47,56). Por otro lado, las huellas T2-L1, TI4 y T7.L1 se caracterizan por presentar una débil mesaxonomía, es decir, el grado en que el dígito III sobresale hacia delante (Lull, 1904). Gracias a ello, se supo que pertenecían a la familia Eubrontiae, y a los primeros terópodos que vivieron en el Cretácico conocidos como Jialingpus. Por otro lado, con los datos de las velocidades de estos organismos (Alexander, 1976) y las longitudes de sus pasos, es posible saber si estaba caminando, corriendo o trotando. Los resultados obtenidos fueron los siguientes: la velocidad de las huellas T1, T2, T4 y T6 fue de 3,96-6.66 km/h, y las longitud del paso de 1,54-1,75 (Leonardi, 1987), lo que implica que estaba caminando; mientras que T5 tiene un valor más alto y una mayor longitud en el paso, por lo que estaba más cerca del trote.
 |
| Figura 4. Huellas de terópodos y pterosaurios. |
Las siguientes
características que tienen en común estas huellas les permitieron identificar el
organismo que las originó: el dígito II aparece representado como una pequeña impresión situado
más próximo al dígito III; el dígito IV es más largo que el III; y el ángulo
divergente entre los dígitos III y IV es de 37º, y eran huellas didáctilas.
Todo ello les sirvió para saber que se trataban de huellas realizadas por Deinonychosaurian. El icnotaxón Deinonychosaurian comprende cuatro icnogéneros Velociraptorichnus, Dromaeopodus, Dromaeosauripus y Menglopipus, que clasifican a las huellas encontradas en tres clases dependiendo de sus tamaños (Figura 5).
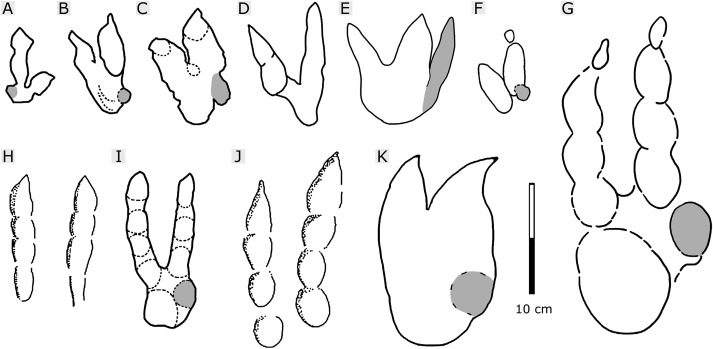 |
Figura 5. Conjunto de dibujos interpretativos de las distintas huellas realizadas por varios icnotaxones de dromeosaurios:
A: Menglongipus (Xing et.al 2009); B: Velociraptorichnus from Shandong (Li et.al 2007); C: Velociraptorichnus from Emei (Xing et. al 2009); D: Velociraptorichnus from Mujiaowu (Xing et.al 2015); E: Velociraptorichnus wangi (Xing et.al 2015); F: Velociraptorichnus encontrado en SMG; G: Dromaeopodus shandongensis (Li et.al 2007); H: Dromaeosauripus jinjuensis (Kim et.al 2012); I: Dromaeosauripus yongjingensis (Xing et.al 2013); J: Dromaeosauripus hamanensis (Kim et.al 2008); K: Dromaeosauripus Jishan (Xing et.al 2013).
|
EL SEGUNDO ESTUDIO: HUELLAS DE SAURÓPODOS.
Como ya he mencionado anteriormente, estas huellas no fueron
las únicas que encontraron. Descubrieron un camino de tres huellas muy bien
conservadas bastante anchas, que nombraron como SMG-S1-S3, en los bloques 1 (figura 3), bloque 2 (figura 7) , bloque 5 (figura 8 y 9) y bloque 6 (figura 6).
Tanto las huellas
SMG-S1 y SMG-S2 tenían una anchura media (WAP/P'ML) de 1.3 la y 1.2 respectivamente. En cuanto a las impresiones del pes eran más largas que anchas; presentaban un
alto grado de heteropodia; y los dígitos del I al IV disminuían de tamaño. Tras
analizarlas, se dieron cuenta de que pertenecían al icnogénero Brontopodus (Lockley et.al 1995), (Lockley el.al 2002).
 |
| Figura 6. Mapa de huellas encontradas en el Bloque 6. En la imagen real de la derecha aparece un camino de huellas realizado por un dromeosaurio. |
 |
| Figura 7. Dibujo lineal de un segmento del camino de huellas del Bloque 2 en el que se muestra la traza del pes bien definidos. |
 |
| Figura 8. Mapa de huellas identificadas en el Bloque 5. |
 |
| Figura 9. Conjunto de huellas realizadas por saurópodos encontradas en el bloque 5. |
SMG-S3 muestra muestra un patrón de marcha inusual, debido a que solo conversan las impresiones derechas del manus, y se encuentran rotadas 21º hacia afuera desde el eje del camino de huellas. También se puede observar como el borde distal de la huella RM2 y el borde proximal de RM3 se encuentran deformadas, cuya posible causa puede ser que se haya formado posteriormente a la huella LP3. (figura 7).
Pero de repente, se dieron cuenta de que les faltaban las impresiones izquierdas del manus. ¿Qué podía haber ocurrido? ¿Cuál ha podido ser la causa?.

Las posibles hipótesis con respecto a la ausencia de impresiones izquierdas del manus podrían ser las siguientes: Un solapamiento producido por otras huellas (Xing et.al 2013); causas litológicas; causas patológicas (McCrea et.al 2015), como una lesión o enfermedad.Se calculó la velocidad del saurópodo que originó estas huellas y se dieron cuenta de que eran las mismas que la SMG 1-2, por lo que se pudo descartar la opción del modo de andar tripedal causado por una lesión.
EL TERCER ESTUDIO: HUELLAS DE ORNITÓPODOS.
Un camino de huellas bastante abundante en todos los bloques
encontrados, excepto en el bloque 6, era producido por otro organismo diferente
a los anteriores. Esta vez se trataba de huellas realizadas por ornitópodos,
caracterizados por impresiones tridáctilas del pes que carecen de almohadillas
en las falanges, aunque a veces se encuentran divididos por unos pliegues
interdigitales adquiriendo una configuración cuatripartita, con tres dígitos y
una almohadilla en el talón, y con ausencia de impresión de manus.
 |
| Figura 8. Conjunto de huellas principalmente de ornitópodos encontradas en el Bloque 4. |
 |
| Figura 9. Conjunto de huellas encontradas en el Bloque 3 realizadas por ornitópodos bípedos con una huella de saurópodos superpuesta (marcada por la flecha). |
Finalmente,
concluyeron que estas huellas pertenecían a la icnofamilia Iguanodontopodidae
(Vialov 1988), pero se encontraron con un pequeño problema:

¿A qué icnoespecie podrían pertenecer?
Para ello, realizaron una tabla
comparativa en el que analizaron cada tipo de huella y las posibles
icnoespecies. Dudaban entre Carirchnium magnificum (Leonardi et.al 1984), Caririchnium leonardii (Lockley 1987), Caririchnium protohadrosaurichnos (Lee 1997), Caririchnium lotus (Xing et.al 2007), Caririchnium
kyoungsookimi (Lim et.al 2012), Caririchnium encontrado en
Shimiaogou, y Ornithopodichnus masanensis (Kim et.al 2009).
 |
| Figura 10. Dibujos interpretativos de huellas realizadas por: A: Caririchnium leonardii; B: Caririchnium protohadrosaurichnos; C: Caririchnium protohadrosaurichnos; D: Caririchnium lotus; E: Caririchnium kyoungsookimi; F: Caririchnium encontrado en Shimiaogou; G: Ornithopodichnus masanensis. |
 Tras un intensivo estudio...
Tras un intensivo estudio...
Comprobaron que no concordaban del todo bien las huellas encontradas con los dibujos interpretativos que se aprecian en la figura 10. Por lo tanto, tenían los suficientes argumentos para crear una nueva icnoespecie, a la que llamaron Caririchnium liucixini (Leonardi et.al 2009).
 |
| Figura 11. Imágenes tomadas de las huellas encontradas en los Bloques 7 y 8, junto a su mapa interpretativo. El Bloque 7 contiene huellas realizas por la icnoespecie Caririchnium liucixini (SMG-O9-R1-L1). |
EL CUARTO ESTUDIO: HUELLAS DE PTEROSAURIOS.
Cuando
pensaban que el estudio había finalizado, les quedaban aún por analizar las huellas
de pterosaurios, que aparecen en las figuras 4 y 6. Decidieron que
pertenecían a estos organismos por las siguientes características: la impresión
plantígrada y tetradáctil del pes es estrecha; tienen un talón en forma de U;
el ángulo de bifurcación entre los dígitos I y III es muy alto, de 131º; y
aparecen las impresiones de tres dígitos I-III. Por lo tanto, concluyeron que
pertenecen al icnogénero Paraechinus (Xing et.al 2013).
LLEGÓ LA HORA DE LA DESPEDIDA...
Y aquí finaliza mi tercera entrada. Espero que no os haya parecido aburrido y hayáis estado entretenidos leyéndolo. Odio las despedidas, pero espero con entusiasmo poder escribiros de nuevo queridos lectores.
¡HASTA LA PRÓXIMA!

REFERENCIAS:
LLEGÓ LA HORA DE LA DESPEDIDA...
Y aquí finaliza mi tercera entrada. Espero que no os haya parecido aburrido y hayáis estado entretenidos leyéndolo. Odio las despedidas, pero espero con entusiasmo poder escribiros de nuevo queridos lectores.
¡HASTA LA PRÓXIMA!

REFERENCIAS:
- Alexander, R.M. 1976. Estimates of speeds of dinosaurs. Nature, 261: 129-130.
- Kim, J.Y., Lockley, M.G., Kim, H.M., Lim, J.D., Kim, S.H., Lee, S.J., Woo, J.O., Park, H.J., Kim, H.S. & Kim, K.S. 2009. New Dinosaur Tracks from Korea, Ornithopodichnus masanensis ichnogen. et ichnosp. nov. (Jindong Formation, Lower Cretaceous): implications for polarities in ornithopod foot morphology. Cretaceous Research, 30: 1387-1397.
- Leonardi, G. 1987. Glossary and Manual of Tetrapod Footprint Palaeoichnology. Departamento Nacional de Producao Mineral, Brazil, p. 75.
- Lim, J.D., Lockley, M.G. & Kong, D.Y. 2012. The trackway of a quadrupedal ornithopod from the Jindong Formation (Cretaceous) of Korea. Ichnos, 19: 101-104.
- Lockley, M.G., 1987. Dinosaur footprints from the Dakota Group of Eastern Colorado. Mountain Geologist, 24: 107-122.
- Lockley, M.G., Hunt, A.P. & Meyer, C.A., 1994b. Vertebrate Tracks and the Ichnofacies Concept: implications for Paleoecology and Palichnostratigraphy. In: Donovan, S. (Ed.), The Paleobiology of Trace Fossils. Wiley and Sons, Inc, New York, pp. 241-268.
- Lockley, M.G., Wright, J., White, D., Li, J.J., Feng, L. & Li, H. 2002. The first sauropod trackways from China. Cretaceous Research, 23: 363-381.
- Lull, R.S. 1904. Fossil footprints of the Jura-Trias of North America. Memoirs of the BostonSociety of Natural History 5: 461-557.
- McCrea, R.T., Tanke, D.H., Buckley, L.G., Lockley, M.G., Farlow, J.O., Xing, L.D., Matthews, N.A., Helm, C.W., George Pemberton, S. & Breithaupt, B.H. 2015. Vertebrate ichnopathology: pathologies inferred from dinosaur tracks and trackways from the Mesozoic. Ichnos, 22: 3-4, 235-260.
- Olsen, P.E. 1980. A comparison of the vertebrate assemblages from the Newark and Hartford Basins (Early Mesozic, Newark Supergroup) of Eastern North America. In: Jacobs, L.L. (Ed.), Aspects of Vertebrate History, Essays in Honor of Edwin Harris Colbert, pp. 35-53.
- Xing, L.D.,Wang, F.P., Pan, S.G. & Chen,W. 2007. The Discovery of Dinosaur Footprints from the Middle Cretaceous Jiaguan Formation of Qijiang County, Chongqing City. Acta Geologica Sinica (Chineseedition) 81 (11): 1591-1602.
- Xing, L.D., Lockley, M.G., Pi~nuela, L., Zhang, J.P., Klein, H., Li, D.Q., Wang, F.P., 2013a. Pterosaur trackways from the Lower Cretaceous Jiaguan Formation (BarremianeAlbian) of Qijiang, Southwest China. Palaeogeography, Palaeoclimatology, Palaeoecology, 392: 177-185.
- Xing, L.D., Lockley, M.G., Zhang, J.P., Klein, H., Marty, D., Peng, G.Z., Ye, Y., McCrea, R.T., Persons, W.S.I.V. & Xu, T. 2015a. The longest theropod trackway from East Asia, and a diverse sauropod-, theropod-, and ornithopod-track assemblage from the Lower Cretaceous Jiaguan Formation, southwest China. Cretaceous Research, 56: 345-362.
- Zhen, S., Li, J., Zhang, B., 1994. Dinosaur and bird footprints from the Lower Cretaceous of Emei County, Sichuan, China. Memoirsof the Beijing Natural History Museum, 54: 105-120.
1 comentario:
Hola Paula, me ha resultado bastante sorprendente la enorme cantidad de información que se puede extraer de los icnofósiles. Yo personalmente, pensaba que la información que se podía extraer de una simple huella, no iba más allá de reconocer el tipo de animal del que provenía, de saber el tamaño del animal que lo produjo, o de si dicho animal se encontraba caminando o corriendo (que ya es mucho). Sin embargo, me he dado cuenta que la icnología va mucho más allá, y por ejemplo, en el artículo de Dánae de las huellas de Tirannosaurus (“Siguiendo a Tiranosaurus paso a paso”), mediante fórmulas, se puede llegara a conocer la velocidad exacta a la que se desplazaba el animal, o como hemos leído en este artículo, se puede saber la especie exacta o incluso reconocer ¡nuevas especies a partir de las huellas!.
Leyendo este artículo, me he dado cuenta que cualquier muestra, por insignificante que parezca (como huellas, minúsculos fragmentes de hueso, etc), nos pueden aportar una gran cantidad de información, y que muchas veces ni nos esperamos. ¡Así es la ciencia!
Publicar un comentario