
Las entradas que he realizado en este blog han
tratado sobre reptiles de un periodo especifico Cretácico y de su
identificación bien por vertebras o por dientes.
He querido que estas entradas trataran de reptiles
dado que me parecen muy importantes ya que supusieron la auténtica colonización
del medio terrestre.
En el Carbonífero superior un orden de anfibios (Anthracosauria) dio origen a los
reptiles, en concreto a Captorrinomorfos, y esto supuso la colonización del medio
terrestre ya que mostraban numerosas adaptaciones para evitar la deshidratación
que supone la vida en un ambiente fuera del agua. Estas características son: un
ventrículo parcialmente separado, una epidermis con escamas para evitar la
evaporación del agua corporal, conductos excretor y reproductor independientes
y un huevo protegido por cáscara (amniota).
La complejidad del huevo amniota sugiere que la
transición entre los anfibios y los reptiles no tuvo lugar más que una vez, y
que por lo tanto el grado evolutivo de los reptiles es monofilético. Así el
huevo amniota se convirtió en la característica evolutiva más importante de los
reptiles y constaba de:
Una membrana protectora calcificada en cuyo interior
una serie de membranas alimentan y protegen al embrión. El embrión está
conectado a un saco vitelino (un saco de sustancias de reserva de las que se
alimenta el embrión durante su estancia en el interior del huevo) y a una
cavidad alantoidea (donde se acumulan las sustancias de desecho en forma de
precipitados nitrogenados). A su vez la membrana amniótica, que cubre la
cavidad amniótica (donde se sitúa el embrión), lo protege del contacto de la
cascara cubriéndolo.
Este huevo supuso la total independencia del medio
acuático por parte de los reptiles, ya que la cavidad amniótica rellena de
líquido semeja el ambiente acuático necesario para el desarrollo del embrión.
Para clasificar las nuevas líneas evolutivas que
surgieron en los reptiles amniotas, científicos realizaron una clasificación
basándose en el número de ventanas o aberturas en la región temporal del
cráneo. Según esta clasificación hay cuatro tipos de cráneos:
 |
| Figura 1. Tipos de cráneos de reptiles amniotas basada en el número de ventanas en la región temporal el cráneo. |
Según este carácter el árbol evolutivo de los
amniotas sería:
 |
|
Figura 2. Árbol evolutivo de los amniotas. En rojo
están los grupos a los que pertenecen los reptiles que he tratado en mis otras
entradas.
|
Como se ve en la figura 2, todos los amniotas, tanto
los extintos como los que aún viven se agrupan en dos grupos iniciales:
-
Synapsida: que dieron lugar a los
mamíferos.
-
Sauropsida: los reptiles propiamente
dichos.
En esta entrada nos vamos a centrar en los Diapsida
y más en concreto en los órdenes de reptiles mencionados en mis otras entradas.
Plesiosauria:
El orden Plesiosauria (plesiosaurios) consiste en
dos superfamilias. El Pliosauroidea (pliosaurios) son típicamente de cuello
corto, mientras que Plesiosauroidea (plesiosaurios) son típicamente de cuello
largo. Sin embargo, las generalizaciones basadas en la longitud del cuello se
consideran poco fiables ya que la investigación sobre la evolución y filogenia
plesiosaurio sigue revelando una historia más complicada. La familia
Polycotylidae, por ejemplo, es un grupo del Cretácico Tardío de plesiosaurios
de cuello corto que se consideraron tradicionalmente como pliosaurios. Sin
embargo, hay evidencias craneales (Carpenter, 1997) que apuntan a una afinidad
plesiosauroidea para ellos. Del mismo modo, los géneros de pliosauroidae Eurycleidus y Attenborosaurus son pliosaurios con cuellos relativamente largos.
 |
|
Figura 3. Esqueleto de Meyerasaurus victor en vista ventral (inferior) (de Smith [2008],
redibujado de Fraas [1910]).
|
Plesiosauria se caracteriza por la presencia de un
conjunto de caracteres derivados y adaptaciones secundarias extremas a la vida
en el agua. Todos los plesiosaurios tienen cuatro grandes aletas y cuerpos
generales rígidos. El esqueleto postcraneal de plesiosaurios se puede dividir
en el esqueleto axial (la columna vertebral y las costillas) y el esqueleto
paraxial (cinturas de las extremidades y las extremidades). Todos ellos poseen
las siguientes sinapomorfias (caracteres derivados compartidos) en su esqueleto
(Rieppel, 1997; Carroll, 1988; Storrs, 1993):
-
Ventralmente expandido (alargado)
cinturas pectoral y pélvicas. Las cinturas pectorales se expanden en placas
delgadas ventrales, el principal elemento de la cintura pectoral son los
coracoides y la escápula siempre que éstos cumplan en la superficie lateral
(exterior) que forman una cavidad glenoidea para acomodar la extremidad
anterior. La escápula puede o no puede cumplir en la línea media dependiendo de
la especie. Un gran complejo de clavícula-interclavícula está presente en los
primeros plesiosaurios, pero se pierde en formas posteriores.
La cintura pélvica al igual que en
la cintura pectoral los elementos forman placas expandidas. El pubis está
situado por delante y el isquion posteriormente. El ilion es un pequeño
elemento que sirve de puente entre la cintura escapular y las costillas sacras
y sólo contacta con el isquion en plesiosaurios.
 |
| Figura 4. Cintura pectoral, A: anterior, y B de lado. |
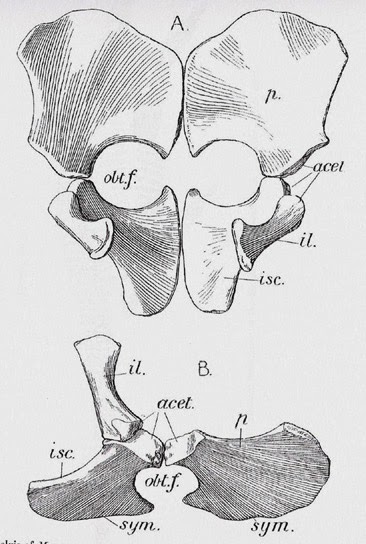 |
| Figura 5. Cintura pélvica, A: anterior y B: de lado. |
-
Ausencia de huesos nasales.
-
Ausencia de contacto entre ilion y pubis.
-
Tronco relativamente corto (cuerpo) y la
cola.
-
Gastralia muy bien desarrolladas y
posicionadas entre la cintura pectoral y pélvica es una cesta de apretadas
costillas ventrales o gastralia. Cada fila se compone de un elemento central en
forma de boomerang, y cerca de tres simples gastralia en cada lado.
-
Acortamiento y ensanchamiento de huesos
de las extremidades e hiperfalangia: tanto el primer plano y las patas traseras
se han convertido en aletas en forma de alas, y se parecen mucho entre sí. El
propodial (húmero / fémur) es relativamente largo, los epipodiales (radio,
cúbito / tibia, peroné) se acortan, haciéndose más anchos que largos en algunos
taxones. Los mesopodiales son en forma de disco. Los metapodiales y falanges
(huesos de los dedos) son bastante cortos y entallados. Muestran hiperfalangia
(aumento en el número de huesos en los dígitos). Puede o no puede haber más o
menos número de huesos accesorios en la zona epi-mesopodial, dependiendo de la
especie.
 |
|
Figura 6. Extremidades vistas desde
abajo, A: extremidad posterior y B: extremidad anterior.
|
-
Dos agujeros pequeños subcentrales
nutritivos (aberturas pequeñas, presumiblemente para los vasos sanguíneos o los
nervios) en la superficie ventral del centrum (una excepción: Brachauchenius y los
agujeros reducidos en algunos otros pliosaurios).
Squamata:
Son un grupo de gran éxito, esta familia de
reptiles incluye lagartos (por ejemplo, gekkota, Escincos, camaleones),
serpientes y anfisbenas. Con más de 7.000 especies, están presentes en todos
los continentes excepto en la Antártida, y han invadido en ambientes marinos
(por ejemplo, serpientes de mar, Mosasaurios), así como la diversificación en muchas
formas terrestres especializadas diferentes, incluyendo animales de madriguera
(por ejemplo anfisbenas), planeadores (por ejemplo, Draco), los corredores
bípedos (por ejemplo, Basiliscus), los escaladores (como los camaleones) y
depredadores activos (por ejemplo Varanus).
Además de la condición de reptil ovivíparo, algunos
grupos dan a luz a crías vivas (vivíparos). Lo más sorprendente es que hay al
menos ocho grupos que contienen una especie con sólo miembros femeninos, que se
reproducen por partenogénesis.
Squamata es el grupo hermano de Rhynchocephalia
(Tuatara y parientes extintos), compartiendo un ancestro común alrededor de
hace 250 millones de años al comienzo del Mesozoico. Juntos, forman el grupo
monofilético Lepidosauria.
Sinapomorfias:
Kinesis craneal (un alto grado de flexibilidad entre
los huesos de la parte posterior del cráneo, que permite movimientos relativos
entre ellos).
Hemipenes apareados (presentes en todos los lepidosaurios) son totalmente eversible.
Dentición pleurodont (dientes fijados en el lado de las superficies interiores de las mandíbulas, y reemplazados periódicamente).
Pérdida de gastralia (costillas ventrales del vientre).
Quinto metatarsiano de doble gancho, funcionalmente análogo al talón de los mamíferos.
Complejidad de la articulación mesotarsal (en comparación con lepidosaurios no squamata).
Hemipenes apareados (presentes en todos los lepidosaurios) son totalmente eversible.
Dentición pleurodont (dientes fijados en el lado de las superficies interiores de las mandíbulas, y reemplazados periódicamente).
Pérdida de gastralia (costillas ventrales del vientre).
Quinto metatarsiano de doble gancho, funcionalmente análogo al talón de los mamíferos.
Complejidad de la articulación mesotarsal (en comparación con lepidosaurios no squamata).
Características:
Debido a que vivían en
las zonas secas, desarrollaron escamas y placas para protegerse de la
desecación. Ellos desarrollaron gruesas capas de la epidermis y de la dermis con el fin de protegerse a sí mismos. La epidermis es una estructura uniforme de material muerto que ya no puede crecer; lagartos y serpientes cambian de piel periódicamente.
La coloración y patrón de los lagartos y las serpientes se determina por el tipo y la disposición de las células de pigmento que son controladas por los nervios o las hormonas. El color o patrón de un lagarto y serpiente cambia cuando se está ocultando o realizando un ritmo diario de actividad.
 |
| Figura 7. Cráneos de Lepidosauria. Articulaciones del cráneo y la mandíbula (flechas) ayudan a distinguir los miembros del orden Squamata (lagartos y serpientes) el uno del otro y de los miembros del orden estrechamente relacionado Rhynchocephalia. Cráneo de un Tuatara (a), un miembro de la orden Rhynchocephalia, es relativamente rígida. Los pivotes de la mandíbula inferior del extremo inferior del hueso cuadrado (verde) y el hueso quadratojugal adyacente. En cráneos de squamata, incluyendo los camaleones (b), la mandíbula pivota desde el extremo inferior del hueso cuadrado. El extremo superior del hueso cuadrado también disfruta de una cierta libertad de movimiento, una condición conocida como streptostyly, por lo que es posible que el animal mueva la parte posterior de la mandíbula inferior más libremente en la manipulación de presa. Muchos squamata tienen una articulación adicional en el cráneo que permite flexionar el hocico hacia arriba y hacia abajo. En la mayoría de los lagartos (c), esa articulación está detrás de los ojos (mesokinesis). Pero en las serpientes (d), esta articulación se encuentra en frente de los ojos (prokinesis). Las serpientes tienen otras articulaciones del cráneo y de la mandíbula flexibles; la víbora (d), por ejemplo, también puede rotar sus huesos de la mandíbula superior que tienen los colmillos. |
Una regresión en el arco del lóbulo temporal
proporciona espacio para los músculos más grandes.
Las dos primeras vértebras en la parte de atrás (eje y Atlas) se modifican de tal manera para hacer posible un movimiento de rotación del cráneo. Para poder ver bien el entorno y defenderse de los depredadores.
Las dos primeras vértebras en la parte de atrás (eje y Atlas) se modifican de tal manera para hacer posible un movimiento de rotación del cráneo. Para poder ver bien el entorno y defenderse de los depredadores.
El estribo y los huesos individuales en los tímpanos reconocen bajas
frecuencias. Por lo tanto, los lagartos y las serpientes eran capaces de oír
los depredadores y sus presas.
Los dientes están anquilosados o fijados por el tejido calcificado. Los dientes están fijos en la parte superior de una cresta ósea en la mandíbula. A lo que se llama dentición acrodont.
La cintura pélvica consta de tres elementos vinculados: ilion, pubis e isquion.
El íleon se conecta en un extremo a dos vértebras sacras por costillas y en el otro extremo de dos elementos de la cintura ventral, pubis e isquio, por tejido conectivo fibroso.
Los dientes están anquilosados o fijados por el tejido calcificado. Los dientes están fijos en la parte superior de una cresta ósea en la mandíbula. A lo que se llama dentición acrodont.
La cintura pélvica consta de tres elementos vinculados: ilion, pubis e isquion.
El íleon se conecta en un extremo a dos vértebras sacras por costillas y en el otro extremo de dos elementos de la cintura ventral, pubis e isquio, por tejido conectivo fibroso.
Saurischia:
La característica principal de Saurischia es la
pelvis, pero también tiene otras. Los saurisquios tienen la pelvis como la de
los lagartos, lo cual es diferente a la pelvis de aves ornitisquios.
Saurisquios incluye a muchos dinosaurios, incluyendo los saurópodos y los
terópodos.
La pelvis tiene la
sínfisis mirando hacia la parte delantera del animal, a diferencia de los
ornitisquios, que tienen el pubis señalando hacia la parte posterior del animal
como un pájaro. |
|
Figura 8. Pelvis de Saurischia y Ornitischia.
|
Sauropoda eran grandes herbívoros como Apatosaurus y
Diplodocus. Theropoda eran carnívoros bípedos, como Compsognathus (tamaño de
una gallina), Deinonychus, Velociraptor, Dilophosaurus y Tyrannosaurus.
Los dinosaurios saurisquios tienen un conjunto de
características únicas. Las siguientes son las características básicas, a parte
de la pelvis, de un dinosaurio Saurischia:
-
Herbívoros.
-
Cabezas pequeñas con forma de espátula o
los dientes en forma de lápiz para rasgar bien las plantas.
-
Por lo general, caminaron a cuatro
patas, pero pudieron extenderse hasta dos con el fin de alcanzar el follaje.
Los terópodos aparecieron por primera vez durante el
Triásico tardío hace unos 230 millones de años (Ma) y se incluyeron como los
únicos grandes carnívoros terrestres del Jurásico Temprano por lo menos hasta
el final del Cretácico, hace unos 65 Ma. En el Jurásico, las aves evolucionaron
de pequeños terópodos especializados coelurosauria, y están hoy representados
por 9.900 especies vivas. Entre las características que unen los dinosaurios
terópodos con las aves son los pies de tres dedos, una espoleta, huesos llenos
de aire, ovivíparos y (en algunos casos) plumas.
Si bien históricamente se generalizada a los terópodos como carnívoros,
de hecho muestran una amplia gama de dietas. Todos los hallazgos iniciales de
fósiles de terópodos mostraron que eran principalmente carnívoros. Especímenes
terópodos conocidos por los científicos en los siglos 19 y principios del 20
todos mostraban afilados dientes con bordes dentados para cortar carne, y
algunos ejemplares aún mostraron evidencia directa de la práctica abusiva. Por
ejemplo, un fósil de Compsognathus fue encontrado con un lagarto en su
estómago, y una muestra de Velociraptor se encontró encerrado en combate con un
Protoceratops.
Los primeros terópodos no carnívoros encontrados
fueron therizinosaurs, originalmente conocidos como segnosáuridos.
Therizinosaurs poseían grandes abdómenes para la elaboración de alimentos de
las plantas, y las pequeñas cabezas con pico y dientes en forma de hoja.
Continuando el estudio de los terópodos maniraptora y sus relaciones mostró que
therizinosaurs no fueron el único miembro de este grupo que no fuera carnívoro.
Varios otros linajes de maniraptora muestran adaptaciones para una dieta
omnívora.
Crocodilia:
Las 22 especies de cocodrilos existentes son
notablemente similares. Diversos detalles morfológicos han sido utilizados para
subdividir en tres familias, Alligatoridae (los caimanes, Caiman, Palaeosuchus, Melanosuchus y caimanes Alligator), Crocodylidae (Crocodylus y Osteolaemus cocodrilo enano) y el Gavialidae (el gavial Gavialis y
Falso gavial Tomistoma) cuyas afinidades son inciertos dentro y fuera del
grupo. Australia tiene dos especies de cocodrilos, de agua salada o cocodrilo
de estuario, Crocodylus porosus y
cocodrilo de agua dulce, Crocodylus
johnstoni.
Los cocodrilos son estudiados como reptiles con
forma de lagarto, con cabeza y tronco bien blindados y con la piel y escamas
córneas, reforzadas con placas óseas. En Crocodylia modernos, la cabeza es
generalmente aplanada y el hocico alargado. Las cápsulas nasales se encuentran
cerca de la punta de la boca y las fosas nasales que puede cerrar en las
narinas externas tienden a elevarse por encima de la punta de la boca; así sumergidos
son poco visibles cuando cazan a presas terrestres. Los
miembros de Alligatoridae tienden a tener hocicos grandes, mientras que los de
la Crocodylidae van desde amplio a largo y delgado. En gaviali el hocico es
extremadamente delgado.
El esqueleto de cocodrilo es típico de la de los
tetrápodos en general y de los archosaurios, en particular, con el cráneo y la
pelvis particularmente especializados. También de interés son la estructura de
las vértebras de la cabeza (atlas y axis), los osteodermos, y la gastralia, o
costillas de estómago, que protegen la superficie ventral.
Como archosaurios, los cocodrilos son diápsidos,
aunque la ventana post-temporal se reduce. La ventana palatal y mandibular
externa y los latero-esfenoides osificados están presentes. La pared de la caja
craneana está osificada, pero supra-temporales y post-frontales están ausentes.
El pterigoideo y hueso cuadrado se adhieren ajustados a la pared lateral de la
caja craneana y la muesca ótica cierra posteriormente. Todas las especies carecen
de agujero parietal. El premaxilar, se amplía postero-dorsalmente, aislando los
fosas nasales externas de los maxilares. Fosas nasales internas se prolongan
posteriormente por un paladar secundario que se forma de los procesos palatinos
de los premaxilares, maxilares, palatinos y pterigoideos y las coanas
secundarias abiertas dentro de los pterigoideos.
La piel de los cocodrilos es gruesa y córnea, y está
revestido de escamas no solapadas conocidas como scutes, dispuestas en filas y
patrones regulares. Estas escamas están siendo producidas por la división
celular en la capa subyacente de la epidermis, el estrato germinativo, y la
superficie de los escudos individuales sacude periódicamente. La superficie
exterior de los escudos se compone de la relativamente rígida beta-queratina,
mientras que la región bisagra entre los escudos contiene sólo la más flexible
alfa-queratina.
Muchos de los escamas se ven reforzadas por placas
óseas conocidas como osteodermos, que son del mismo tamaño y forma que las
escamas superficiales pero crecen por debajo de ellas. Estas son las más
numerosas en la espalda y el cuello del animal y pueden formar una armadura
protectora. A menudo tienen, crestas, bultos prominentes y están cubiertos en
beta-queratina resistente. La mayor parte de la piel de la cabeza se fusiona
con el cráneo. La piel en el cuello y los flancos está suelta, mientras que en
el abdomen y la parte inferior de la cola están revestidas en grandes escamas cuadradas
dispuestas en filas ordenadas.
Bibliografía y referencias:
Carpenter, K.
1997. Comparative cranial anatomy of two North American Cretaceous Plesiosaurs.
191-216. In Callaway, J. M and Nicholls, E. L. (eds.). Ancient Marine
Reptiles. Academic press. London. 501pp.
Fraas, E. 1910. Plesiosaurier
aus dem oberen Lias von Holzmaden. Palaeontographica, 57, 105-140.
Rieppel, O. 1997.
Part 2: Sauropterygia. Introduction. 107-119. In Callaway, J. M and
Nicholls, E.L. (eds). Ancient Marine Reptiles. Academic press. London.
501pp.
Storrs, G. W.
1993. Function and phylogeny in sauropterygian (Diapsida) evolution. American
Journal of Science. 293-A. 63-90.
Trabajos utilizados:
Phil R. Bell,
Federico Fanti, Mark T. Mitchell and Philip J. Currie (2014) Marine Reptiles (Plesiosauria and Mosasauridae) from
the Puskwaskau Formation(Santonian-Campanian), West-Central Alberta. Journal of
paleontology, V. 88 nº 1 p. 187-194.
Funston, G. F.
& Currie, P. J. (2014). A previously undescribed caenagnathid mandible from
the late Campanian of Alberta, and insights into the diet of Chirostenotes
pergracilis (Dinosauria: Oviraptorosauria). Canadian Journal of Earth Sciences, 51(2): 156-165.
No hay comentarios:
Publicar un comentario